


A Grand Unified Theory of Consciousness
How associative learning spurred the evolution of consciousness

There is a widespread belief that a biological explanation of consciousness is impossible because an unbridgeable explanatory gap exists between the science of neurobiology and our conscious experience.1 On this way of thinking, nothing that we know about the biology of cognition can account for my rich subjective experience of the smell of grass in the morning or the taste of an apple. An influential version of this skepticism comes from David Chalmers, who believes that consciousness has no physical cause but is rather a fundamental non-physical property. Others, the “mysterians,” do not deny that consciousness has a biological cause, but believe that its workings and origins are so mysterious that humans are incapable of understanding it.
There has never been any good reason to believe in Chlamers’ theory. It is highly unlikely that physicists have missed some fundamental property of the universe that will turn out to account for consciousness. However, mysterianism has greater credibility. Neurobiologists have explained much about the workings of different components of consciousness, like perception and emotion. However, until recently, none of them had articulated a coherent and convincing total biological theory of consciousness that could explain how the different components interacted, how consciousness evolved, and how we can distinguish between conscious and non-conscious animals. The absence of such an overarching theory has made consciousness seem mysterious indeed.
Now, however, we have the first serious candidate for a grand unified biological theory of consciousness that can bridge the explanatory gap. In their 2019 book, The Evolution of the Sensitive Soul: Learning and the Origin of Consciousness, Israeli biologists Simona Ginsburg and Eva Jablonka (G&J) propose that consciousness evolved to enable what they call “unlimited associative learning” (UAL) and that understanding this type of learning is the key to answering the deepest questions about the nature of consciousness. Certainly, much of the neurobiology underlying consciousness remains unexplained, but G&J provide a compelling framework for future research.
Unlimited Associative Learning
In my last essay, I discussed the nature and evolution of limited associative learning (LAL), a simple form of learning that preceded UAL. LAL involves associating a simple stimulus with an innate reflex. I discussed the example of a sea slug learning to associate a light touch to its siphon with a gill withdrawal reflex. An animal capable of UAL, by contrast, can associate a compound stimulus, like a red, one-foot wide, rubbery ball with a compound motor action consisting of multiple steps that is learned rather than encoded in the animal’s genome. Compound stimuli can also consist of processes that unfold over time, like a melody or a sequence movements.
If a pigeon learns to peck at a red button to obtain a food reward, then this is an example of LAL, as the pigeon associates a simple stimulus, the color red, with the innate reflex of pecking. However, if a pigeon learns to associate a food reward with pecking three times on a picture of a tomato plant, then two times on the picture of a face, this is an example of UAL, as the pigeon is performing a compound learned sequence of actions and recognizing compound objects.
Associative learning evolved because it conferred fitness benefits on its possessors. Learning animals can respond more flexibly to their environments, which gives them an advantage in predation and escaping predation, finding food, finding mates, and so forth.
Unlimited Associative Learning as the Origin of Consciousness
G&J’s argument is that LAL does not require consciousness, but UAL does, and that consciousness evolved to make UAL possible. To make this argument, G&J assemble a list of common characteristics of consciousness from the literature on the subject and argue that UAL plausibly spurred their evolution. Among the defining traits of consciousness are the capacities for:
The binding and the unification of percepts into compound stimulus
Global brain activity (the processes underlying consciousness are distributed throughout the brain)
Attention, behavioral and cognitive plasticity, and learning
Temporal thickness (percepts persist in working memory)
Values, emotions, and goals
Embodiment and selfhood (the animal understands its difference from the external world and has a distinctive perspective on the world)
The binding of percepts into compound stimuli is part of the definition of UAL. Simple sensory neurons are sufficient to guide the behavior of animals incapable of UAL. However, sensory integration areas that associate different percepts into compound stimuli are necessary for UAL. Similarly, motor integration areas are required for compound actions.
Compound stimuli often consist of quite complex patterns in the world and sequences that unfold over time, so discerning them is not always an instantaneous process. This discernment may require a period of investigation and interpretation, which requires an animal to store information about the object in short-term or working memory. Working memory also expands an animal’s capacity for associative learning, as it allows an animal to associate events that do not occur at the same time. From the development of working memory comes the temporal thickness characteristic of consciousness.
As I noted in my last essay, associative learning requires an animal to distinguish between associations that are important and those that can be safely ignored. If an animal cannot do this, then overlearning becomes a problem. This problem becomes especially acute with the development of UAL, which expands the range of what an animal can sense along with the number of possible associations it can form. Avoiding irrelevant stimuli requires hierarchical predictive processing that generates prediction errors. The brain must form a model of the world that tells the animal what to expect so that it can avoid getting distracted by the predictable and focus on the surprising and novel elements of its environment. UAL requires an interplay between bottom-up and top-down sensory impulses whereby expectations are compared with reality.
A neurological example of this interplay between bottom-up and top-down processing is vision. The brain has many different visual processing areas that work together to create vision. Different areas are specialized for different stages in visual processing. V1 processes lines, V4 processes shapes, and the inferotemporal cortex processes complex objects. The flow of information through these regions is not one-directional but bi-directional. Higher levels of the visual system are constantly interpreting and shaping the input from lower ones.
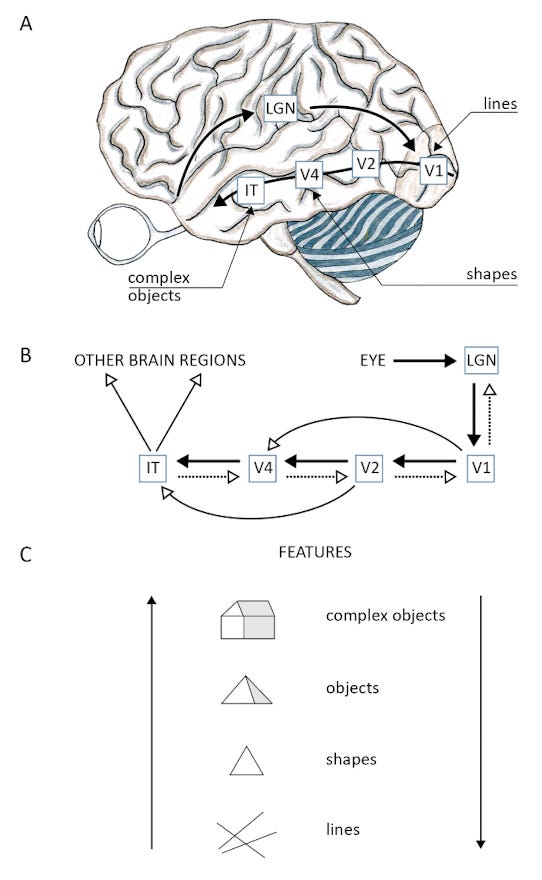
When the animal’s model of the world does not match reality, a prediction error is generated. Prediction error leads to the development of attention, which is a top-down signal to select the surprising stimulus for focus. This focusing attentiveness enables cognitive and behavioral plasticity or the capacity to learn new behaviors.
Another mechanism that helps the animal to focus on relevant stimuli is a flexible evaluation system. UAL spurs the evolution of the senses of pleasure and displeasure, which function as a common currency that allows the ranking of different options. Which food the animal pursues is determined by which food brings it greater pleasure. If an animal sees appetizing food, but also smells a predator in the area, it can make a decision about what to do by weighing the pleasure of eating against the displeasure of risking death. In this way, UAL requires emotions for successful functioning.
As we saw in the last essay, a sense of self is necessary for any kind of associative learning, as the animal must be able to distinguish the effects of its own movements from those of external beings. If an animal uncovers food through its exploratory efforts, it must know that it itself is responsible for finding the food to learn from the event. With UAL comes a new form of selfhood. The wealth of options for learning opened up by UAL causes animals to have distinctive life histories, knowledge, and perspectives. Many animals capable of UAL can exhibit idiosyncratic behaviors and personalities.
For all of these types of cognition to work, global brain activity is required. UAL spurs the development of more powerful sensory organs and larger and more variegated brains to handle the problems of sensory and motor integration, predictive processing, and flexible value systems. Naturally, all of this equipment has to work in synchrony to produce UAL.2
Further support for G&J’s thesis comes from the fact that, at least as far as scientists know, all animals that are capable of UAL are also capable of the characteristics that define consciousness.
Neuroanatomy is further evidence for G&J’s theory. All animals capable of UAL also have relatively large and variegated brains with areas that are specialized for the functions that make consciousness possible. These animals have brain areas that are responsible for processing sensory information into compound stimuli, assembling compound motor actions, forming memories, and evaluating sensory input. G&J conclude that consciousness is present in almost all vertebrates, many arthropods (e.g. honeybees and fruit flies), and some mollusks (e.g. octopus and cuttlefish).
UAL leads to the development of rich mental representations of the world that are lacking in animals incapable of this type of learning. A conscious mental representation is characterized not only by sensory detail and object and pattern recognition, but also by subjective experience. Perceived objects are understood in the context of an animal’s past learning and value system so that each stimulus has a unique essence for a unique being. G&J name conscious mental representations “categorizing sensory states” (CSSs):
These representational sensory states...categorize, through their dynamics, both input and output. The inputs that elicit the mental representational capacities activate memory traces of other inputs of the same type (for example, inputs and memory traces related to hunger), and these inputs and traces also determine what type of response will occur as memory traces of the motor responses to the inputs and the stimuli associated with the attainment of the attractor (relief from hunger) become active. At the same time, particular CSSs are highly differentiated, since each CSS always maps a specific input-output relation. (p. 375)
These CSSs are the biological basis of rich subjective experience, and they bridge the explanatory gap between biology and consciousness.
The Evolution of Unlimited Associative Learning
G&J believe that UAL first appeared in the Cambrian Era, which began 542 million years ago and lasted around 60 million years. This era saw a rapid development and diversification of new species called the “Cambrian Explosion.” Before the Cambrian, only the simplest animal nervous systems existed. The Cambrian saw the evolution of all currently existing animal phyla and of the first brains.
G&J argue that the evolution of associative learning was one of the main drivers of this flowering of life. The sensory capacities of animals also evolved rapidly: the first compound eyes, antennae, and tentacles evolved at this time. These sensory organs provided the inputs that enabled new types of complex, learned, and intelligent behavior, including learning-based predation and escape from predation, active swimming using fins and flaps, walking on the sea floor, burrowing, brood caring, mating displays, and physical competition over mates.
While insects, birds, and mice are probably conscious, human consciousness is quite different from theirs. In his 1996 book Kinds of Minds, Philosopher Daniel Dennett proposed that the evolution of cognition takes place in four stages.
Darwinian organisms are incapable of learning, and their behavior consists entirely of innate reflexes. These include single-celled microorganisms, plants, fungi, and the simplest animals, like sponges.
Skinnerian organisms can adapt their behavior to different conditions through associative learning. These are named after the psychologist and theorist of associative learning, B.F. Skinner.
Popperian organisms can imagine different behaviors and outcomes and opt for one as opposed to others. They are named after philosopher Karl Popper, who said that foresight permits our hypotheses to die in our stead.
Gregorian organisms, named after psychologist Richard Gregory, can use language and culture to augment their intelligence. While rudiments of language and culture exist in other species, humans are the only truly Gregorian creatures.
The Evolution of the Sensitive Soul is primarily about the evolution and nature of Skinnerian organisms. Humans have cognitive abilities that no other organism is capable of, so our conscious experience is undoubtedly quite different from that of other animals.
Can zombies exist?
G&J formulate a plausible and empirically supported theory of how consciousness works and how and why it evolved based on psychology, biology, neuroscience, and paleontology. I find the theory exciting because it possesses the combination of simplicity and massive explanatory power that is one of the hallmarks of great scientific theories.
However, the philosopher David Chalmers, who has many fans on Substack, thinks that theories like G&J’s are ill-conceived. He believes that no physical facts can explain consciousness, and that it is, therefore, impossible to explain consciousness through biology. He believes that other aspects of human cognition, like learning and the mechanics of perception, can be illuminated by biology, but that “the impressive progress of the physical and cognitive sciences has not shed significant light on the question of how and why cognitive functioning is accompanied by conscious experience” (25). (Chalmers has not revised his opinion since stating it in his 1996 book, The Conscious Mind.) Rather, Chalmers believes that consciousness is due to a non-physical property of the universe.
Chalmers’ theory depends on the premise that consciousness and learning are separable activities. In his famous zombie thought experiment, he asks us to consider whether it is logically coherent to imagine a zombie who is an exact copy of a person but has no consciousness. He specifies that this zombie could behave just like the original. The premise on which his entire philosophy of consciousness is founded is that the concept of a zombie is logically coherent.
Chalmers’ ideas have met with a variety of criticisms, but G&J’s ideas make one of these criticisms particularly compelling: a non-conscious exact replica of a human being is impossible because consciousness is essential to human learning and cognition. Non-conscious creatures like sea slugs are capable only of the most rudimentary kind of cognition. It is indeed logically impossible for a zombie to mimic a human being when it does not have one of the abilities that would make this mimicry possible.
For this reason and others, G&J consider Chalmers’ notion that consciousness needs a non-biological explanation “profoundly wrong” (p. 453). I hope to see future research that builds on G&J’s grand unified theory of consciousness. However, it seems time to admit that the theories like Chalmers’ have reached a dead end.
I have substantially changed this essay for the sake of clarity since I first posted it.
G&J formulate simplified models of how UAL works. In chart below, they present a model of classical conditioning by which an animal learns to associate one percept with another and respond to that sequence with a motor output or learned behavior. For example, a dog owner usually feeds their dog at 5:30 PM every day when they return from work. The dog learns to associate the sounds, sights, and smells of its owner returning with getting fed. Sometimes the owner forgets, so the dog learns to tap its bowl with its paw and bark to remind them.
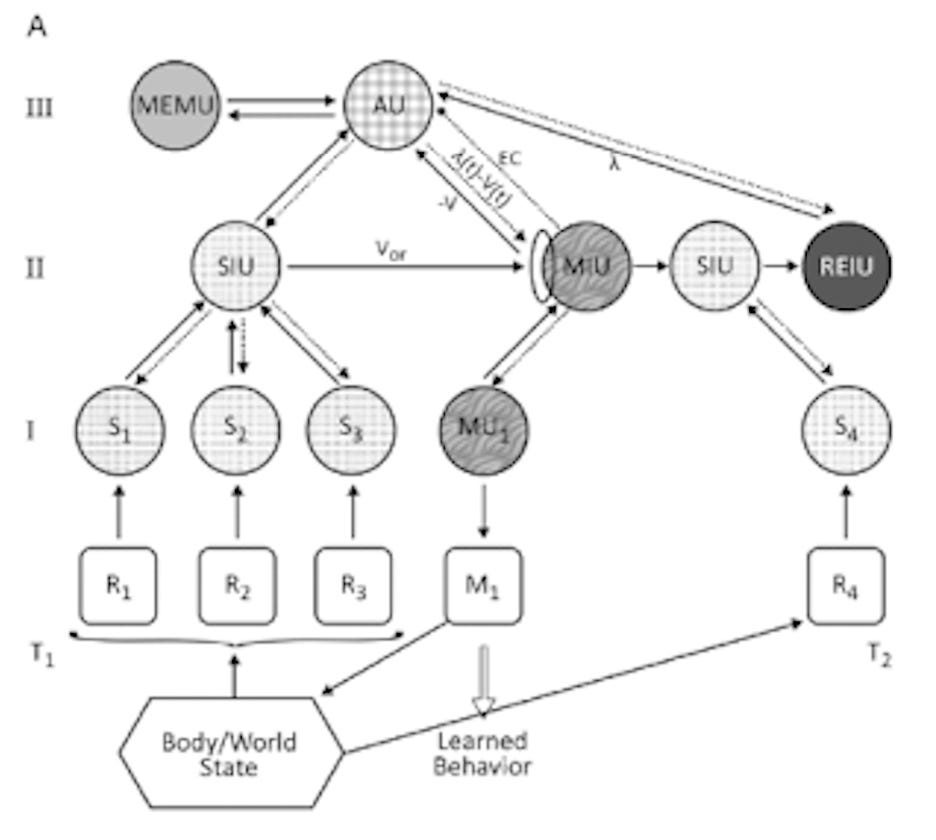
In this chart, at Time 1 (T1), an animal perceives an object through multiple sensory receptors (R1 etc.), which pass them on to sensory processors (S1 etc.). These are integrated into a compound stimulus by a sensory integration unit (SIU): the owner is returning home. Not only does information flow from the sensory receptors to the SIU, but information from the SIU, as well as the memory unit (MEMU), flows down to the sensory units. This top-down information represents the animal’s model of the world, which it integrates with sensory input. At Time 2 (T2), a stimulus (the food) is received by the reinforcing unit (REIU), which enables any percept to be reinforced. The REIU sends the association unit (AU) information about the reward strength, λ. If the reward is different from that predicted by the animal’s model of the world, a prediction error is generated. By this process, the dog learns that the owner’s return is important because it is associated with the surprising reward of food. The prediction error is represented by the math feeding into the ellipse next to the motor integration unit (MIU). The prediction error is based on the discrepancy signal: PE = λ(t) − V(t), where the received reward is λ, and the reward prediction at time T1 is V. The connection between the stimulus in the SIU and the MIU is strengthened or weakened based on the size of the prediction error. After the dog learns the association between the owner returning and food, the owner’s return stimulates the motor output of going to the bowl and tapping it. The learned information gets encoded in the memory unit.
Excellent piece, Ian! Thanks for the introduction to Ginsberg & Jablonka. I heavily endorse your concluding statement. Of course, I think it’s almost a rite-de-passage if one is thinking and writing about consciousness to tackle Chalmers and the Mary’s Room sleight of hand, so I can’t resist doing it (planned next post) even though I agree it’s long past it’s best before date.
re octopus: tiles are just flat oyster shells.